4 June 2026
The views and opinions expressed in this article are solely those of the original author(s) and do not necessarily represent those of GARDP, their donors and partners, or other collaborators and contributors. GARDP is not responsible for the content of external sites.
In the complex war against antimicrobial resistance (AMR), few adversaries are as challenging and as geographically pervasive as the New Delhi Metallo-β-lactamase (NDM). Since its first identification in 2008, this enzyme has become a symbol of the “superbug” crisis in the media.1,2 NDM-producing bacteria render carbapenems ineffective, drastically limiting therapeutic use in Enterobacterales and other Gram-negative pathogens. NDMs dissemination is predominantly plasmid-borne. The blaNDM gene is commonly linked to mobile genetic elements that favor gene transfer among bacteria. This gene is frequently associated with additional resistance genes to other families of antibiotics, such as fluoroquinolones and aminoglycosides, further limiting treatment options.3 Originally identified in the Indian subcontinent, NDM variants have been reported in different species of bacteria around the globe. A recent CDC report disclosed a 460% increase in NDM-producing Enterobacterales infections in the US between 2019 and 2023.4
NDMs belongs to the family of Metallo-β-lactamases (MBLs), zinc-dependent enzymes able to cleave the β-lactam ring of penicillins, cephalosporins and carbapenems, i.e. virtually all β-lactam antibiotics.5,6 In contrast, monobactams such as aztreonam cannot be hydrolyzed by MBLs. There are three main families of clinically relevant and plasmid-encoded MBLs: Imipenemases (IMPs), Verona Integron-encoded (VIMs) and NDMs. They all share a similar protein fold and zinc binding active site, featured by a shallow and wide active site groove that enables binding of most β-lactam antibiotics. This geometry enables MBLs to bind newly designed antibiotics with bulky substituents that may not fit into the more restricted active site cavity of serine-β-lactamases (SBLs) such as those from the KPC, AmpC, OXA and CTX-M families.5,6 MBLs and SBLs have different catalytic mechanisms: as a result, they are not inhibited by the available SBLs inhibitors. β-lactam-based inhibitors, such as clavulanic acid, are hydrolyzed by MBLs but do not give rise to an inhibited adduct. Instead, avibactam binds poorly and is not hydrolyzed by MBLs (see below). This results in a broader resistance phenotype conferred by MBLs compared to SBLs. However, this does not account for the larger prevalence in the last decade of NDMs over proteins from the VIM and IMP families.
Are NDMs unique among MBLs?
A distinctive feature of the NDM family is that all variants are lipidated proteins, anchored to the inner leaflet of the outer-membrane of Gram-negative bacteria, unlike VIM and IMP enzymes, that are soluble, periplasmic proteins.7 This localization enhances the stability of NDMs under the zinc limitation stress imposed by the nutritional immunity response from the mammalian host. Membrane-binding also favors incorporation of active NDMs into outer membrane vesicles, that act as shuttles that can transport resistance elements beyond the producing cell.7 NDM-loaded vesicles can protect populations of neighboring bacteria otherwise susceptible to antibiotics, creating a communal resistance phenotype. This vesicle-mediated defense extends the effective range of NDMs beyond the producing cell.7
In the complex war against antimicrobial resistance (AMR), few adversaries are as challenging and as geographically pervasive as the New Delhi Metallo-β-lactamase (NDM).
Until recently, NDMs were characterized by a substrate profile similar to VIM and IMP enzymes, including cephalosporins. However, NDMs are the only MBLs reported to confer resistance against the last-resort cephalosporin-siderophore antibiotic cefiderocol,8 as discussed below.
NDM diversity: why variants matter
The first NDM-1 enzyme was identified in a carbapenem-resistant Klebsiella pneumoniae isolate recovered in Sweden in January 2008 from a patient hospitalized in New Delhi, India.9 Since then, we have witnessed the emergence and spread of 93 NDM variants.10 This is a large number compared to the documented 111 IMP variants (since 1988) and 96 VIM variants (since 1999).
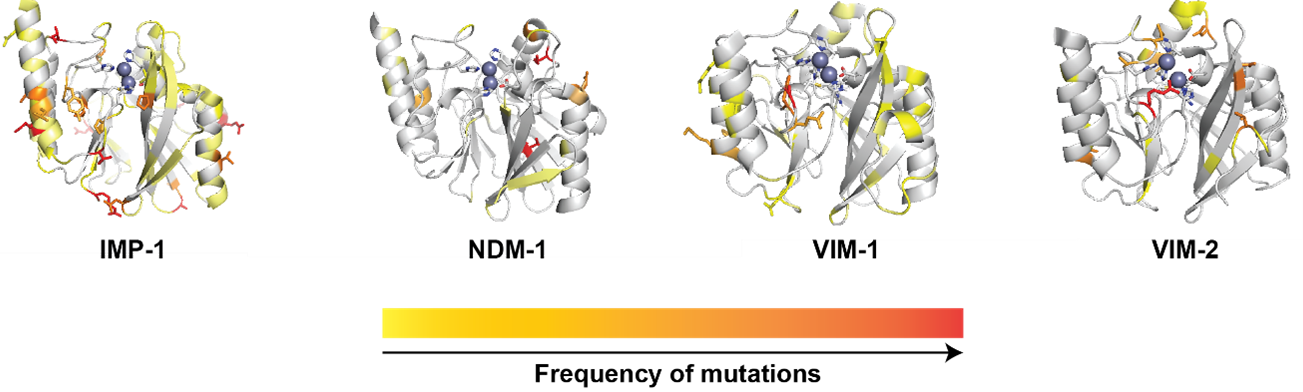
Figure 1: Structures of IMP-1 (PDB 4C1G), NDM-1 (PDB 4EXY), VIM-1 (PDB 5N5G) and VIM-2 (PDB 1KO3) highlighting the positions presenting sequence variation within the enzyme groups. The positions are colored in the cartoon representation according to increasing absolute frequency of mutation within enzymes belonging to each group.
NDM variants typically differ by a small number (one to three) of substitutions, often outside the active site (Figure 1). In contrast, the IMP and VIM families show a more diversified phylogenetic tree, with up to 60 substitutions, mostly in active site loops and second-shell residues, i.e., directly connected to the active site, and therefore shaping their substrate preferences. This is not common in the NDM family, accumulating substitutions outside the active site, which have been shown to have little impact on catalytic efficiency in NDM variants 1 to 8.11 This raises the question of the evolutionary advantage of so many NDM variants.
Most NDM variants are able to provide a better resistance phenotype to different bacterial hosts than NDM-1 under zinc limitation conditions, either by enhancing the zinc binding capability or by stabilizing the zinc-depleted NDM in the bacterial periplasm, preventing their aggregation or degradation.12,13 This is indeed the case of NDM-5, NDM-7 and NDM-9, which are becoming the most prevalent variants after NDM-1.14 In addition to the advantage of being membrane-bound proteins, the accumulation of different substitutions favors the activity of NDMs under zinc-limiting conditions. Therefore, NDM variants are being selected to overcome the impact of zinc starvation at the infection sites. This represents a new paradigm in the evolution of β-lactamases.
Treatment options
There are two possible approaches to treat infections caused by MBL producers within a β-lactam-based strategy: (1) use a β-lactam substrate that can escape hydrolysis from MBLs, or (2) resort to combination therapy using an antibiotic with an MBLs inhibitor.
Aztreonam, a monobactam antibiotic approved in 1986, is not hydrolyzed by any MBL, and targets the penicillin binding protein PBP3 from Gram-negative bacteria. However, MBLs are usually co-expressed with at least one SBL, that can inactivate aztreonam. Therefore, the combination of aztreonam with avibactam, a SBL inhibitor with no activity against MBLs, represents one alternative to treat NDM-producing Enterobacterales. This combination has recently been approved for clinical use15 and represents an alternative to the combination of ceftazidime/avibactam with the addition of aztreonam.16 Ceftazidime targets also other PBPs, therefore potentiating the activity of aztreonam but is susceptible to hydrolysis by MBLs.17
A distinctive feature of the NDM family is that all variants are lipidated proteins, anchored to the inner leaflet of the outer-membrane of Gram-negative bacteria, unlike VIM and IMP enzymes, that are soluble, periplasmic proteins.
Cefiderocol is a siderophore-modified cephalosporin, that can bind iron ions essential for the bacterial cell and is actively incorporated through the iron transporters at the outer membrane. This “Trojan horse” strategy has provided a successful treatment for Enterobacterales and non-fermentors. Despite initial reports suggesting that cefiderocol was refractory to MBL-mediated hydrolysis,18 recent data show that NDM-1 and NDM-5 (but not VIM-2 and IMP-1) can hydrolyze cefiderocol.19 While successful treatments of infections caused by NDM-producing isolates have been reported, most clinical failures with cefiderocol are specifically associated with the presence of NDM variants, underscoring the impact of NDM producers in the clinics.20
Cefepime/zidebactam (WCK 5222) is a β-lactam/β-lactam “enhancer” pairing that has recently completed Phase 3 clinical evaluation. It targets PBP3 (cefepime) and PBP2 (zidebactam), with zidebactam also imparting β-lactamase inhibitory effects and escaping hydrolysis by MBLs.21
The design of MBL inhibitors has been challenging due to the absence of a covalent intermediate in MBL-mediated β-lactam inactivation, and due to the presence of MBL-like proteins in eukaryotes, thus being prone to off-target effects that can induce toxicity in mammalian cells.22 Two recent boronate-based drugs, taniborbactam23 and xeruborbactam,24 can inhibit both MBLs and SBLs, and are or have been evaluated in combination with different antibiotics. However, a resistant phenotype to taniborbactam has been identified, since a single substitution present in NDM-9 renders this enzyme impervious to inhibition by this new compound.25 Boronate compounds provide a novel, promising approach as future therapeutic options.
Concluding remarks
Despite the recent promising therapeutic progress, NDMs are a family of proteins with large evolutionary power, able to adapt to different bacterial hosts and environmental conditions. Therefore, new therapies should be prescribed with caution to limit the generation of resistance in a short period of time, and with close monitoring of the patient clinical trajectory. In addition, precision diagnostics are required, involving the identification of the MBL family and, in some cases, the allelic variants, to guide the best possible therapeutic option.

Clarisa Parodi obtained her Bachelor of Science degree in Biotechnology from the National University of Rosario (UNR), Argentina, in 2020.
She is currently working towards her PhD under the supervision of Prof. Alejandro J. Vila within the Institute of Molecular and Cellular Biology of Rosario (IBR, CONICET-UNR). Her research focuses on the behavior of NDM-expressing Acinetobacter baumannii under infection-relevant conditions.

Alejandro Vila is Professor of Biophysics at the National University of Rosario (UNR), Argentina and Senior Researcher of the Argentinean National Council of Scientific and Technological Research (CONICET).
His laboratory studies the role of β-lactamases in antimicrobial resistance, encompassing biochemical and mechanistic details, protein evolution, inhibitor design as well as β-lactamase physiology in the periplasm of Gram-negative bacteria. He has published more than 180 papers in leading international scientific journals. His laboratory is a world leader in the biochemical study of metallo-β-lactamases.
Conflict of interest:
The authors declare that they do not have any relationships or affiliations that could be construed as a potential conflict of interest.
Republication:
The Viewpoints on our website are to be read and freely shared by all. If they are republished, the following text should be used: “This Viewpoint was originally published on the REVIVE website revive.gardp.org, an activity of the Global Antibiotic Research & Development Partnership (GARDP).”
References
- Dortet L, Poirel L, Nordmann P. (2014) Worldwide dissemination of the NDM-type carbapenemases in Gram-negative bacteria. Biomed Res Int. 249856. doi:10.1155/2014/249856
- Kumarasamy KK, Toleman MA, Walsh TR, et al. (2010) Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect Dis. doi:10.1016/S1473-3099(10)70143-2
- Acman M, Wang R, van Dorp L, et al. (2022) Role of mobile genetic elements in the global dissemination of the carbapenem resistance gene bla NDM. Nat Commun. doi:10.1038/s41467-022-28819-2
- Rankin DA, Stahl A, Sabour S, et al. (2025) Changes in Carbapenemase-Producing Carbapenem-Resistant Enterobacterales, 2019 to 2023. Ann Intern Med. doi:10.7326/ANNALS-25-02404
- Palzkill T. (2013) Metallo-β-lactamase structure and function. Ann N Y Acad Sci. 1277(1):91-104. doi:10.1111/j.1749-6632.2012.06796.x
- Bahr G, González LJ, Vila AJ. (2021) Metallo-β-lactamases in the Age of Multidrug Resistance: From Structure and Mechanism to Evolution, Dissemination, and Inhibitor Design. Chem Rev. 121(13):7957-8094. doi:10.1021/acs.chemrev.1c00138
- González LJ, Bahr G, Nakashige TG, Nolan EM, Bonomo RA, Vila AJ. (2016) Membrane anchoring stabilizes and favors secretion of New Delhi metallo-β-lactamase. Nat Chem Biol. 12(7):516-522. doi:10.1038/nchembio.2083
- Warecki BA, Tamma PD, Bonomo RA, Vila AJ. (2025) NDM-driven cefiderocol resistance: effect and therapeutic considerations. Lancet Infect Dis. S1473-3099(25):00477-3. doi:10.1016/S1473-3099(25)00477-3
- Yong D, Toleman MA, Giske CG, et al. (2009) Characterization of a new metallo-β-lactamase gene, bla NDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. doi:10.1128/AAC.00774-09
- Naas T, Oueslati S, Bonnin RA, et al. (2017) Beta-lactamase database (BLDB) – structure and function. J Enzyme Inhib Med Chem. 32(1):917-919. doi:10.1080/14756366.2017.1344235
- Makena A, Brem J, Pfeffer I, et al. (2015) Biochemical characterization of New Delhi metallo-β-lactamase variants reveals differences in protein stability. J Antimicrob Chemother. 70(2):463-469. doi:10.1093/jac/dku403
- Bahr G, Vitor-Horen L, Bethel CR, Bonomo RA, González LJ, Vila AJ. (2018) Clinical Evolution of New Delhi Metallo-β-Lactamase (NDM) Optimizes Resistance under Zn(II) Deprivation. Antimicrob Agents Chemother. 62(1):e01849-17. doi:10.1128/AAC.01849-17
- González LJ, Bahr G, González MM, Bonomo RA, Vila AJ. (2023) In-cell kinetic stability is an essential trait in metallo-β-lactamase evolution. Nat Chem Biol. 19(9):1116-1126. doi:10.1038/s41589-023-01319-0
- Xia C, Yan R, Liu C, et al. (2024) Epidemiological and genomic characteristics of global blaNDM-carrying Escherichia coli. Ann Clin Microbiol Antimicrob. doi:10.1186/s12941-024-00719-x
- Carmeli Y, Cisneros JM, Paul M, et al. (2025) Aztreonam–avibactam versus meropenem for the treatment of serious infections caused by Gram-negative bacteria (REVISIT): a descriptive, multinational, open-label, phase 3, randomised trial. Lancet Infect Dis. doi:10.1016/S1473-3099(24)00499-7
- Marshall S, Hujer AM, Rojas LJ, et al. (2017) Can ceftazidime-avibactam and aztreonam overcome β-lactam resistance conferred by metallo-β-lactamases in Enterobacteriaceae? Antimicrob Agents Chemother. doi:10.1128/AAC.02243-16
- Tamma PD, Lodise TP, Mojica MF, Perez F, Bonomo RA. (2025) Revisiting REVISIT: The Case for Ceftazidime/Avibactam Plus Aztreonam. Clin Infect Dis. doi:10.1093/cid/ciaf239
- Ito A, Kohira N, Bouchillon SK, et al. (2016) In vitro antimicrobial activity of S-649266, a catechol-substituted siderophore cephalosporin, when tested against non-fermenting Gram-negative bacteria. J Antimicrob Chemother. 71(3):670-677. doi:10.1093/jac/dkv402
- Warecki BA, Tomatis PE, Mojica MF, et al. (2025) Cefiderocol “under siege”? Understanding the rise of NDM-mediated resistance to novel agents. Chem Sci. 16(27):12519-12533. doi:10.1039/D5SC02122G
- Simner PJ, Mostafa HH, Bergman Y, et al. (2022) Progressive Development of Cefiderocol Resistance in Escherichia coli during Therapy is Associated with an Increase in blaNDM-5 Copy Number and Gene Expression. Clin Infect Dis. 75(1):47-54. doi:10.1093/cid/ciab888
- Sader HS, Rhomberg PR, Flamm RK, Jones RN, Castanheira M (2017). WCK 5222 (cefepime/zidebactam) antimicrobial activity tested against Gram-negative organisms producing clinically relevant β-lactamases. J Antimicrob Chemother. 72(6):1696-1703. doi:10.1093/jac/dkx050
- Mojica MF, Rossi MA, Vila AJ, Bonomo RA. (2022) The urgent need for metallo-β-lactamase inhibitors: an unattended global threat. Lancet Infect Dis. 22:e28-e34. doi:10.1016/S1473-3099(20)30868-9
- Liu B, Trout REL, Chu GH, et al. (2020) Discovery of Taniborbactam (VNRX-5133): A Broad-Spectrum Serine- And Metallo-β-lactamase Inhibitor for Carbapenem-Resistant Bacterial Infections. J Med Chem. 63(6):2789-2801. doi:10.1021/acs.jmedchem.9b01518
- Hecker SJ, Reddy KR, Lomovskaya O, et al. (2020) Discovery of Cyclic Boronic Acid QPX7728, an Ultrabroad-Spectrum Inhibitor of Serine and Metallo-β-lactamases. J Med Chem. 63(14):7491-7507. doi:10.1021/acs.jmedchem.9b01976
- Le Terrier C, Gruenig V, Fournier C, Nordmann P, Poirel L. (2023) NDM-9 resistance to taniborbactam. Lancet Infect Dis. doi:10.1016/S1473-3099(23)00069-5